The views below and mine and mine alone. I am a professional within the industry of arboriculture and I do not pretend to know everything or even close. Therefore, please do not take my words as if they are fact. You’re entitled to disagree, to any degree – insofar as the disagreement is reasonable. The hyperlinks are for ease to you the reader (please click them). You may find a previous blog post or two of mine useful, as context, before reading this. I have kept the below as brief as I consider possible that will still provide the necessary degree of context and explanation to demonstrate my opinions.
Scope of this post
This post does not dissect or scrutinise the technical elements of BS 5837:2012 Trees in relation to design, demolition and construction – Recommendations. This post does analyse whether this standard is fit for purpose, in the current legal context.
A background to British Standards
British Standards (hereafter ‘standards’) are published by the BSI Group, an organisation that operates under Royal Charter with the defined purpose of producing and publishing standards and providing related services for the ultimate betterment of industry – specifically, “to provide a reliable basis on which common expectations can be shared regarding specific characteristics of a product, service or process“. The key is therefore in the name: standardisation (which facilitates replicability). Adherence to a standard is voluntary.
Focussing on standards, the Royal Charter permits the BSI Group to “set up, sell and distribute standards of quality for goods, services, and management systems and prepare and promote the general adoption of British and international standards and schedules“. Periodically, published standards must be revised, altered, and amended “as experience and circumstances may require“.
The header and introduction of the Royal Charter for the BSI Group.
How standards are made
Standards are developed in collaboration between representatives of the relevant organisations associated with the scope and objective of the standard, adhering to – and being the only standard that must be adhered to (i.e. adherence is not voluntary) – the standard for producing new standards (BS 0:2016 A standard for standards – Principles of standardization).
Development is the responsibility of the relevant technical committee (e.g. B/213 for trees), which will normally oversee more than one standard. The technical committee is comprised of relevant organisations, companies, and individuals that fairly represent the range of professions affected by the standards it oversees. Within the technical committee, there are working groups that will focus on more detailed elements (e.g. the development of a single standard or aspects of it). The technical committee will therefore develop a draft standard that they will then publish for public consultation (a ‘draft for public comment’ or ‘DPC’), which will last for a minimum of 60 days.
The B/213 technical committee page on the BSI Group website.
Following consultation, the feedback will be collated, analysed by the technical committee, and modifications made to the standard so that it is then published – or, in some cases, work is halted, due to significant concerns over the contents of the draft standard. The decision is down to the consensus of the technical committee
Typically, the production of a standard takes 18 months. Throughout, the operations of the technical committee must be transparent and defensible. The result should be a standard that is desirable, practical, and authoritative, provides no exclusive benefit to any party, serves the needs of the relevant community, and is consistent with regulations and legal principles established at the time of publication.
How standards are reviewed
Published standards must be maintained, in order to remain relevant (and for the BSI Group to adhere to its responsibilities as provided by Royal Charter). Therefore, all standards must be reviewed at least every five years, which can be initiated by the BSI Group or the technical committee. The review is undertaken by the technical committee responsible for the standard. Public consultation remains necessary, for the review of a standard. In some cases, the review may result in the standard being withdrawn.
Trees and construction – the standard
The standard for trees in the context of construction (encompassing all RIBA stages) is BS 5837:2012 Trees in relation to design, demolition and construction – Recommendations (hereafter ‘BS 5837’). This standard replaced the 2005 version. Its purpose is to provide “recommendations relating to tree care, with a view to achieving a harmonious and sustainable relationship between new construction/existing structures and their surrounding trees“. To this end, it sets out the recommended process through which trees should be managed in the context of construction, which includes outside of the planning process (e.g. if building something under permitted development). However, it is principally used as part of the planning process where trees are being managed on or adjacent to construction sites where planning permission is required.
The front cover of BS 5837.
BS 5837 – legal developments since its publication
Since the time of the publication of BS 5837 in April 2012, there have been many important legal developments. These include but may not be limited to:
Of these three legal developments, the NPPF is the crucial development, because it states at paragraph 47 that planning applications must (in 99% of cases) be determined in accordance with the relevant planning policies, which includes at national, regional, and neighbourhood level, in addition to the core that is determining applications against local planning policies.
BS 5837, in its role within the planning process (excluding permitted developments) as a standard document, does not make reference to this crucial element (e.g. there is no reference in the Figure 1 flowchart to consulting planning policy). The first major pit-stop for managing trees in the context of development is relevant planning policy that will give a valuable indication of what is and is not acceptable, in planning terms. Therefore, BS 5837 is not fully fit for purpose, in this regard.
The NPPF also states at paragraph 54 that “local planning authorities should consider whether otherwise unacceptable development could be made acceptable through the use of conditions or planning obligations“, and at paragraph 55 “should be kept to a minimum and only imposed where they are necessary, relevant to planning and to the development to be permitted, enforceable, precise and reasonable in all other respects“. For this reason, the Town and Country Planning (Pre-commencement Conditions) Regulations 2018 refined the approach to conditions insofar as local planning authorities must confirm the details of pre-commencement conditions with the applicant prior to imposing the conditions upon them. Specifically, the manner in which Arboricultural Impact Assessments and Arboricultural Method Statements apply within the current framework of BS 5837 now may routinely differ in practice from what is recommended in theory.
These reasons collectively have a significant material effect upon the manner in which trees should be managed and considered, as part of the planning process. BS 5837 in its current form does not consider these effects and therefore it does not “provide a reliable basis on which common expectations can be shared regarding specific characteristics of a product, service or process“.
An excerpt of a local planning policy that requires trees to be protected should they be retained.
So what?
BS 5837 was reviewed in 2017, according to the BSI Group. In this context, it is not now until 2022 that it must be reviewed. Indeed, it can be reviewed before then though this will arise probably only following concern being raised over an insufficient scope.
My opinion is that BS 5837 cannot ‘wait’ another five years before being reviewed, becuase assuming it takes 18 months to produce a new version of BS 5837 then it will not be until 2023/4 that BS 5837 accounts for the significant changes in the planning process. In this context, BS 5837 will not “provide a reliable basis” for over half a decade from now during which time development is still ongoing and ideally should be speeding up to meet government housing targets.
The second part of this cluster of blog posts is one the first of the duo of talks presented by Lynne Boddy. Lynne is a well-known mycologist and researcher and thus, as regards wood-decay fungi, is a good authority from which we can all learn a substantial amount. For a fungi enthusiast such as myself, learning about fungi from one of the best is, in and of itself, very exciting. However, the information presented was equally as exciting, which I shall run through below.
As a slight aside, please watch out for her new book, which is currently being written and should be finished later this year, currently entitled Fungi in Trees. This book will be aimed at the arboriculturist. Lynne is also planning to re-write Fungal Decomposition of Wood, which was a magnum opus co-authored with Alan Rayner.
Fungi invading trees
Primarily, we must accept one core tenet of wood decay: the anatomy of wood has a massive impact upon mycelial networks that sojourn through the wood substrate and selectively metabolise woody cells and their deposits as they go. Indeed, in an ideal world, fungi would gun right for the ray parenchyma, which are incredibly nutritious living cells. However, these cells are very challenging to get to, by virtue of their ‘aliveness’ – living cells in good condition are not easily devoured. Further to this, the sapwood also offers a great for invading fungi, though again – because of the high moisture condition meaning the environment is largely anaerobic (fungi are aerobes and require oxygen to metabolise) – this part of the wood structure is not easily accessed. Of course, the vascular wilts have a better time invading sapwood that is functional, though many fungi will have to bide their time or arrive opportunistically onto and into dysfunctional sapwood if they are to have any means of success in acquiring the treasures within. Thus, is recognising that such living (i.e. conductive and functional) areas of wood are likely just beyond the reach of many fungi, wood-decayers will typically resort to the back-up food source: non-functional xylem vessels within the heartwood or ripewood.
The heart rotters
Now, in accepting this, we come on to perhaps the broadest cohort of wood-decay fungi in living and standing trees (fallen trees have their dysfunctional sapwood metabolised like ravens engulfing a fresh meal) we know of: the heart rotters. These fungi will enter the central wood (i.e. ‘the heart’) through exposed areas of this central column by either sufficiently deep root, stem or branch injury – ultimately, there generally needs to be a continuity of viable ‘heart’ substrate, if any significant degree of colonisation is to take place. Where continuity doesn’t exist, or the fungus finds itself limited to only a certain area, the only manner in which is will typically be able to continue existing is by (1) exiting and finding another host or (2) biding its time and waiting for currently functional sapwood to become incorporated into the heartwood or ripewood, in which it can then spread into – assuming the tree doesn’t lay down defensive barriers that cannot be breached, in order to protect its sapwood, which is in itself a pursuit undertaken to safeguard and hence sustain the high-moisture content of its sapwood (contrary to Shigo’s model, which infers compartmentalisation is largely there with the end in mind of prohibiting fungal succession into the wood). When we look at the ripewood of beech (Fagus sylvatica), we can observe this phenomenon very well – a rosy-coloured ripewood (red heart) with lots of separation lines between instances of decay successions. Ganoderma australe, Ganoderma pfeifferi and Ganoderma resinaceum will afford the most acutely observable examples of this, given their ability to breach such zones by metabolising the phenolic deposits laid down by the tree (this touches upon the idea of a fifth wall in the CODIT model, which will be discussed more later on).
An old Laetiporus sp. on yew (Taxus baccata).
By virtue of the heartwood or ripewood being the least inhospitable (heartwood, in particular, is still often disgustingly harsh, as regards its environment), we can observe immediately some species-specific associations. For instance, the dyer’s mazegill (Phaeolus schweinitzii) will frequent gymnospermous hosts, such as the cedars (Cedrus spp.) and pines (Pinus spp.), whilst chicken of the woods (Laetiporus spp. – often L. sulphureus, though by all means not always, as we will see) will be found often on oak (Quercus spp.), sweet chestnut (Castanea sativa) and yew (Taxus baccata) – it can, indeed, be found on other hosts, as well. On the face value of things, these three tree species have seemingly little in common, though all three have extractive-rich heartwood that this genus can metabolise effectively. In terms of why the genus is referred to and not the exact species, Lynne is of the stance that what we term ‘chicken of the woods’ and default to as L. sulphureus is actually a variety of different species each with their own specialisations – perhaps even down to a specific host tree species (notably for yew, where the chicken species is most pertinently viewed as being a different one).
From a tree management perspective, Lynne then addressed the importance of the heart rotters – what is their impact? Put simply, they change the way in which we view the tree from a safety perspective; as in, when fruiting bodies of heart rotters are identified, if the tree is standing and there exists a target, management considerations are routinely entertained and sometimes the tree is felled. Additionally to this, however, we have other things to appreciate:
heart rotters do a great job at recycling nutrients, which can then be re-assimilated by the tree when they are uptaken back through its roots (including adventitious aerial roots) or the mycorrhizal fungi the roots associate with
the wood qualities produced by heart rotters are ideal as habitat for saproxylic insects and nesting birds
The heart of this beech has been hollowed-out by decay fungi. In the process, before its failure, what conditions did this decay provide for insects and, crucially, what habitat does it provide now?
Latent colonisers
Considered the specialised opportunists, such fungi are present within the sapwood or bark, as propagules (thick-walled resting spores known as chlamydospores). Biding their time until conditions are right, wherein the sapwood becomes dysfunctional through means such as wounding or drought (causing embolism), they are perhaps most acutely observed in the years after drought years where they can trigger the formation of strip cankers and resultant reaction growth by the tree (see the below photo). Thus, this year, in the UK, is one to watch out for, as regards such fungi (it might also explain why Kretzschmaria deusta was so abundant this year, given Ascomycetes love dry conditions, which prevailed last summer).
A strip canker in beech caused by the latent fungus Biscogniauxia nummularia that has induced reaction growth, which can be seen cross-sectionally in the bottom right image).
At this point, a delegate enquired as to whether Massaria disease of plane (Splanchnonema platani), as an Ascomycete, could be prevailing in urban conditions recently, because of dry conditions (such as London, where it was been very severe these past few years but prior to that un-noticed). Discussions continued and Frank Rinn interjected to add his thoughts:
massaria progresses quickly in dry conditions
recent dry summers have allowed for massaria to thus progress very rapidly (killing branches in as little as three months)
the lowering of water tables in cities for the construction of basement levels of buildings has meant that plane trees can no longer tap into groundwater supplies
mature plane trees afford the best conditions for massaria; notably lower lateral branches, which are shaded from the rest of the crown and thus may be most prone to stress
there have been reports since 1903 from Croatia where large plane trees have shed branches and the massaria fungus was termed the “branch-cleaning fungus”
conditions are collectively ideal for massaria to become prevalent, as they stand
Reverting back to latent fungi, Lynne then mentioned that she considers fungi to be latent across a broad variety of trees. For example, the coal fungus (Daldinia concentrica), whilst found most often on ash (Fraxinus excelsior) can be isolated from the sapwood of a great range of different broadleaved tree species in the UK. It is, indeed, only when specific conditions arise that are preferable for this fungus that it begins to create mycelial networks – such conditions might not arise in particular trees, or may arise only after conditions suitable for other fungi have arisen and thus D. concentrica then has no capacity to colonise the substrate. Hence, ash remains the core host of this species, in the current climate. However, for the jelly ear fungus (Auricularia auricula-judae), which is also a latent fungus within the vascular system, having been found largely solely on elder (Sambucus nigra) in the 1950s, it is now found on over 20 host species – this marks a huge increase in host range, prompted perhaps by changing climatic conditions.
Wall V
The CODIT model, offered to us by Shigo, details four walls – as can be seen here. As mentioned by Frank Rinn, Shigo himself was considering the possibility of a fifth wall, though this never ‘made it into’ the model. However, Lynne argues that there is the potential for a fifth one, which hearkens back to what was discussed above, as regards rot within the heart of beech wood).
Specifically, whilst the barrier zone (fourth wall) is a zone laid down at the time of wounding by the vascular cambium, the dynamic responses by the tree that occur in real-time as fungal decay advances constitutes a distinction from this initial barrier. Indeed, as decay advances, living cells within the heartwood or ripewood (they do exist; though through mechanisms not fully appreciated, but thought to be associated with the rays running radially through the wood), in addition to the functional sapwood, will, in order to protect the sapwood and keeps its high-moisture quality intact, will plug woody cells beyond the current zone of decay with extractives and phenolic compounds – this will occur within the sapwood most often, though may also be able to occur in the heartwood around the regions where pockets of living cells exist. This response resultantly produces incredibly dense zones of wood that afford the tree’s sapwood a means of protection, which it would otherwise lack, assuming the barrier zone (Wall IV) failed to contain fungal decay. Of course, if this fifth wall fails, another will form, and so on and so forth.
See the myriad of demarcations across this cross-section of a beech that failed from decay by Ganoderma pfeifferi, which suggest a dynamic fifth wall being effective.
Perhaps we will see this idea discussed more in Lynne’s re-write of Fungal Decomposition of Wood.
Fungal succession
When a tree decays, the fungi that initiated the decay process will not end it. This is because, much like all other ecosystems, as an environment changes those organisms that are best-placed to utilise it change as well. In this sense, fungi are no different – they succeed into a dynamic and ever-altering substrate (wood). Such a phenomenon can be so readily observed when walking into any woodland, when comparing the fungi on standing trees and those in early stages of decay and those much more heavily decomposed. In instances where an entire tree falls and stays largely intact, succession can be most acutely observed, as Lynne detailed with a little help from Ted Green and some research students.
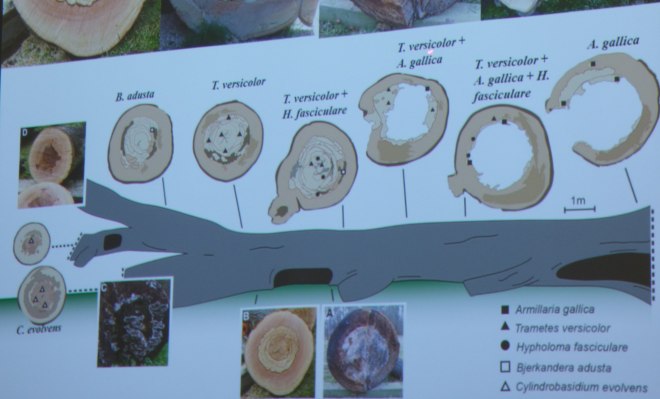
The tree in question, a mature beech, failed and was left, in sections, for ease of its movement into an accessible place, to decay. Along the beech, it was found, through analysis of the wood in the laboratory and by presence of fruiting bodies, different fungi were observed colonising different parts at different stages of decay (as shown below). Such an observation does seem readily apparent, though to have it confirmed through scientific means affords us with an understanding that is more concrete than merely the anecdotal. Indeed, whilst Trametes gibbosa was not isolated from this beech, the presence of Bjerkandera adusta infers that, at some point in the future, T. gibbosa will be found – it parasitises upon the mycelium B. adusta, before then colonising the wood substrate itself. We are, in a sense, therefore, witnessing fungal warfare.
The different fungi found at different parts of the beech at different stages of the decay process (open in a new tab to see this in a slightly larger size).
Delving further into the notion of fungal warfare, what is essentially meant is chemical warfare. Fungi synthesise and secrete enzymes, which they use principally to degrade wood, though that can also be used to defend territory or attack other fungi. The result of any fungal battle can be one of four things:
deadlock, whereby neither fungi gains any ground against the competing fungus
replacement, whereby one fungus loses its territory entirely by the other
partial replacement, whereby one fungus loses of some its territory to the other
mutual replacement, whereby the fungi essentially ‘trade’ places with one another and neither gains any net ground
So how does one determine the outcome of any such skirmish, you ask? Unfortunately, there are so many variables in play that even pitting two fungi against one another in a laboratory is only going to give a slight allusion to what really occurs, though there does nonetheless exist a limited hierarchy of combativeness from which we can assume who the victor will be, under most circumstances (see here at 25:09 timestamp).
Of course, even in assessing this we still have so many caveats to throw in. For example, where moisture conditions are drier because the wood is more exposed, Ascomycetes (i.e. Hypoxylon spp.) will have a better time in securing more wood substrate, as they operate effectively under dry conditions. Indeed, the wood qualities of the substrate itself will even play a role – did the tree uptake pollutants during its life or is it exposed to such pollutants currently, for example. More crucially, if a dead piece of wood (or entire tree) is standing and has thus been subject to relatively dry and exposed conditions suddenly falls to the woodland floor, those fungi reigning when the tree was standing will likely succumb to wood-decay fungi adapted to higher moisture levels and cooler more stable conditions.
The next part of this series will be a brief one on bacteria in wood, as discussed again by Lynne Boddy. I hope to have that written up in the coming few days.
Everything written here is supported by sources I have referenced (check for yourself), as always, so do not treat this as an assault on either side and / or their respective religions. This post is through the lens of the tree, so treat it as such. Moreover, the entire topic is very interesting.
As Israel collides with Palestine, trees are – and always have – been caught up in the melee. Principally, olive and citrus groves, some of which may have been tended to for many centuries by the Palestinians (Temper, 2009), are bulldozed or otherwise uprooted, with little respect for their cultural and historical importance (Allen, 2008; Graham, 2002). As an example, in 1986, when the Israeli military seized Midya, over 3,300 olive trees were uprooted, and a further 2,000 olives were bulldozed in Qattana (Bardenstein, 1999). Some of the trees removed from Qattana were later re-planted within the Jewish sector of West Jerusalem (Lentin, 2000), though by that point the damage (in many an aspect) had certainly been done. Some Israeli residents did protest their planting (out of anger towards the state), by tying ribbons to the trees that contained messages such as “Take me back to Qattana!” (Bardenstein, 1999), whilst others, across the entire conflict, have chained themselves to the olive trees in order to stop the bulldozers from uprooting them (Sfard, 2009), supported Palestinian farmers by helping them harvest from their olive trees and, at times, defending them in the process (Stephan, 2003), provided replacement olive tree for those uprooted (usually by settlers), or helped to retain olive trees within occupied territories for their symbolic meaning of peace (Braverman, 2009) – “extending the olive branch“, per se.
Below the surface level of removing ‘enemy’ trees, the removal of olive trees has a very political undertone. Olive trees have been held in very high regard by Palestinians for generations (and are regarded by some as holy trees), where they were farmed and thus supported viable economies (Braverman, 2009; Cohen, 1993), and their removal (or ‘capture’, by where groves were encompassed into the territory of Israel) by Israeli forces therefore can also be interpreted as an attack on Palestinian culture and custom (Bardenstein, 1999; Bowman, 2007; Braverman, 2009; Kershner, 2005) – notably when such acts are supported by the Court (Sfard, 2009). In some cases, it may even be Jewish settlers who vandalise or cut down the olive trees (Kershner, 2005), and even when the Israeli army have allowed the Palestinians to harvest their olive crops. In such instances, the Israeli army will generally not intervene (Pigni, 2010).
Such a political (and, to a marked degree, religious) act may be most pertinently discerned when the olive groves (or individual trees) are captured or destroyed during harvesting season, which has indeed occurred in some instances (Batniji et al., 2009). Moreover, the fact that many olive groves have been uprooted (comprising of tens of thousands of individual olive trees – in Qafeen alone, 12,600 olives were uprooted for this reason) for the construction of the Separation Barrier in the West Bank was also a cause of huge upset, for the Palestinians; particularly when their uprooting was coupled with justifications including to construct watchtowers, roads, checkpoints, and other security fences (all of which further hamper daily life and privacy), in addition to the use of the groves for sheltering armed Palestinians (Braverman, 2009). For those groves not uprooted, the Separation Barrier may instead have isolated Palestinian farmers from their olive trees, for much of the year. In Qafeen, over 100,000 olive trees suffered this isolated fate.
Olive trees are removed to facilitate the construction of the Separation Barrier. Source: Haaretz.
Whilst the capture and removal of Palestinian groves has been ongoing, Israel has also been afforesting barren regions of its territory – and for many decades. Spearheaded largely by the Jewish National Fund that was established in 1901 (and since 1961 has been Israel’s exclusive forestry agency), the afforestation program was, at its core, a religious, ecological, and territorial pursuit (Amir & Rechtman, 2006; Bardenstein, 1999; Braverman, 2009; De-Shalit, 1995; Ginsberg, 2000; Stemple, 1998; Tal, 2013), with pine species (including Pinus halepensis) being particular favourites (Osem et al., 2008; Weinstein-Evron & Galili, 1985). In recent decades, the emergence of numerous pests associated with the pine (such as Matsucoccus josephi) has however led to more diverse plantations, with other pine species (including Pinus brutia) and deciduous tree species being selected for use (Braverman, 2009).
In essence, a core reason for this afforestation is because Israel, in the ages gone by, was considered to be covered with forests (even up to the 11th century A.D., in places), though it is suggested that when the Jewish people were in exile those who occupied Israel (from around 722 B.C. – 1948 A.D.) destroyed many of these forests (due to arson, harvesting for fuel, overgrazing, sabotage, and warfare) and thus, upon the return of the Jewish people to Israel, in order to bring Israel back to its former character, forests were (and still are) planted upon the barren slopes (Stemple, 1998; Tal, 2012; Tal, 2013). Braverman (2009) states that the Jewish National Fund has planted over 200,000,000 trees across more than 225,000 acres of claimed land, since its inception. However, according to the Old Testament, in the book of Joshua, even Jewish peoples have been responsible for some of this historic clearance in their Promised Land (Tal, 2013), and for this reason the Jewish National Fund is seeking to restore Israel’s forests of ten thousand years ago – soon after the last glacial ice age. In fact, a great deal of planting, each year, is undertaken in the leading up to – and on the day of – Tu B’shvat (Bardstein, 1999; Zerubavel, 2000).
Hiran Forest in 1998 (left) and 2008 (right). Source: KKL-JNF.
After the creation of the Jewish National Fund, though prior to its major afforestation practices towards the middle of the century, the British had, since 1918 (after they had seized southern Palestine), planted up many hundreds of thousands (if not many millions) of saplings (comprising of species including stone pine, tamarisk, terebinth, and oaks) on the hills of Israel (Tal, 2013), and before this (from 1860 onwards) the Turkish Ottoman Empire and settling German Templars had done much the same (Ginsberg, 2006; Liphschitz & Biger, 2004).
In this afforestation project, such planted areas are also oft designated as forest reserves and thereby protected by Israeli law, which Braverman (2009) dubs as “lawfare” against the Palestinians, whose land may have been afforested following seizure. This planting up of forest on occupied lands, of which a sizeable portion was planted over destroyed Palestinian villages in the years after 1948 (an act of camouflage, and for some allegedly the camouflage of war crimes), also makes the land very difficult to reclaim, as the reclaimers must first remove all of the trees (after gaining the permission to clear the perhaps protected forest); in this sense, Palestinians may never be able to occupy such land again, be it for living within or for cultivation. In some cases, Braverman (2009) writes, Palestinians have even retaliated against this afforestation by firing rockets into the planted pine forests or burning the pine forests through arson, with a desire much aligned to Israel’s uprooting of the olive trees (in a sense, a ‘tree for a tree’). In this respect the tree, and specifically the pine, is a tool of war, and thus represents the enemy as a solider would (Boerner, 2011; Braverman, 2008).
On a more philosophical level, the fact that the Jewish National Fund would plant a tree for each newborn from Jerusalem in Jerusalem’s artificially-borne Peace Forest, dedicate the specific tree to the child, and provide the individual with a certificate (including a photo of the tree) that remarks on how it is hoped the tree and child grow together, outlines the innate affinity (or interchangeability) man has with trees (Braverman, 2009); as is detailed before this blog post on earlier ones associated with trees and religion.
Furthermore, the populist and globally crowd-funded nature of a fair portion of the tree planting, supported via financial gifts (complete with material rewards, such as memorial stones) and the use of the ‘Blue Box’ (located in households, schools, and offices), sewed into the fabric of the afforestation project a very emotionally evocative and inclusive aspect to both children and adults of the Jewish faith, even if the donator was geographically separated from Israel (Bar-Gal, 2003; Braverman, 2009; Zerubavel, 2000). Perhaps, this ability for a Jewish person to fund the planting of a tree may dampen their feeling of loss for not living within the Promised Land; in place of their presence, they can fund the planting of a tree, which can be considered a “proxy immigrant” (Braverman, 2009). At a tangent, the returning of the landscape to forest is also important on a cultural level, because the forests were incredibly important for the Jewish peoples’ ancestors; often would children be named after trees, and even Israel itself was sometimes compared to a tree (Zerubavel, 2000; Zerubavel, 2005).
References
Allen, L. (2008) Getting by the occupation: How violence became normal during the Second Palestinian Intifada. Cultural Anthropology. 23 (3). p453-487.
Amir, S. & Rechtman, O. (2006) The development of forest policy in Israel in the 20th century: implications for the future. Forest Policy and Economics. 8 (1). p35-51.
Bar-Gal, Y. (2003) Propaganda and Zionist Education: The Jewish National Fund, 1924-1947. USA: University of Rochester Press.
Bardenstein, C. (1999) Trees, forests, and the shaping of Palestinian and Israeli collective memory. In Bal, M., Crewe, J., & Spitzer, L. (eds.) Acts of Memory: Cultural Recall in the Present. USA: University Press of New England.
Batniji, R., Rabaia, Y., Nguyen–Gillham, V., Giacaman, R., Sarraj, E., Punamaki, R., Saab, H., & Boyce, W. (2009) Health as human security in the occupied Palestinian territory. The Lancet. 373 (9669). p1133-1143.
Boerner, R. (2011) Trees as soldiers in a landscape war. Landscape Ecology. 26 (6). p893-894.
Bowman, G. (2007) Israel’s wall and the logic of encystation: Sovereign exception or wild sovereignty?. Focaal. 50 (1). p127-135.
Braverman, I. (2008) “The Tree Is the Enemy Soldier”: A Sociolegal Making of War Landscapes in the Occupied West Bank. Law & Society Review. 42 (3). p449-482.
Braverman, I. (2009) Planted Flags: Trees, Land, and Law in Israel/Palestine. USA: Cambridge University Press.
Cohen, S. (1993) The politics of planting: Israeli-Palestinian competition for control of land in the Jerusalem periphery. USA: University of Chicago Press.
De‐Shalit, A. (1995) From the political to the objective: the dialectics of Zionism and the environment. Environmental Politics. 4 (1). p70-87.
Ginsberg, P. (2000) Afforestation in Israel: a source of social goods and services. Journal of Forestry. 98 (3). p32-36.
Ginsberg, P. (2006) Restoring biodiversity to pine afforestations in Israel. Journal for Nature Conservation. 14 (3). p207-216.
Graham, S. (2002) Bulldozers and bombs: the latest Palestinian–Israeli conflict as asymmetric urbicide. Antipode. 34 (4). p642-649.
Kershner, I. (2005) Barrier: the seam of the Israeli-Palestinian conflict. USA: Palgrave Macmillan.
Lentin, R. (2000) Israel and the Daughters of the Shoah: Reoccupying the Territories of Silence. USA: Berghahn Books.
Liphschitz, N. & Biger, G. (2004) Green Dress for a Country – Afforestation in Eretz Israel: The first hundred years 1850-1950. Israel: KKL.
Osem, Y., Ginsberg, P., Tauber, I., Atzmon, N., & Perevolotsky, A. (2008) Sustainable management of Mediterranean planted coniferous forests: an Israeli definition. Journal of Forestry. 106 (1). p38-46.
Pigni, A. (2010) A first-person account of using mindfulness as a therapeutic tool in the Palestinian Territories. Journal of Child and Family Studies. 19 (2). p152-156.
Sfard, M. (2009) The Price of Internal Legal Opposition to Human Rights Abuses. Journal of Human Rights Practice. 1 (1). p37-50.
Stemple, J. (1998) Viewpoint: a brief review of afforestation efforts in Israel. Rangelands. 20 (2). p15-18.
Stephan, M. (2003) People power in the Holy Land: How popular nonviolent struggle can transform the Israeli-Palestinian conflict. Journal of Public and International Affairs. 14 (Spring). p164-183.
Tal, A. (2012) Israel’s New Bible of Forestry and the Pursuit of Sustainable Dryland Afforestation. Geography Research Forum. 32 (1). p149-167.
Tal, A. (2013) All the Trees of the Forest: Israel’s Woodlands from the Bible to the Present. USA: Yale University Press.
Temper, L. (2009) Creating facts on the ground: Agriculture in Israel and Palestine (1882-2000). Historia Agraria. 48 (1). p75-110.
Weinstein-Evron, M. & Galili, E. (1985) Prehistory and paleoenvironments of submerged sites along the Carmel coast of Israel. Paleorient. 11 (1). p37-52.
Zerubavel, Y. (2000) The Forests as a National Icon: Literature, Politics, and the Archeology of Memory. In Elon, A., Hyman, N., & Waskow, A. (eds.) Trees, Earth, and Torah: A Tu B’Shvat Anthology. USA: The Jewish Publication Society.
Zerubavel, Y. (2005) The forest as a national icon: literature, politics, and the archaeology of memory. Israel Studies. 1 (1). p60-99.
The arthropods are vast in terms of species, and include ants, beetles, butterflies, mites, moths, spiders, and so on. Therefore, covering the entire spectrum of arthropods in this section is impractical, though the general provisioning by trees will be outlined and species will be used to illustrate given examples.
Many arthropods are considered to be saproxylic in nature – they principally utilise dead woody material (both standing and fallen, in both dead and living trees) as habitat, for at least part of their life cycle, though they may also rely upon fungal sporophores associated with the presence of deadwood, as is to be detailed below (Gibb et al., 2006; Harding & Rose, 1986; Komonen et al., 2000). Of all the saproxylic arthropods, beetles are perhaps the most significant in terms of the proportion occupied of total saproxylic species worldwide (Müller et al., 2010), though saproxylic flies also feature in great numerical abundance (Falk, 2014; Harding & Rose, 1986).
Beetles may be either generalist or specialist in nature (on either broadleaved or coniferous hosts), and they will normally require a host with an abundance of deadwood (or large sections of coarse woody debris) usually over 7.5cm in diameter that resides within an area typically not heavily shaded (Müller et al., 2010; Siitonen & Ranius, 2015). This may be, in part, due to many beetle species (in their adult stage) requiring nectar from herbaceous plants, which would be lacking in woodland with significant canopy closure (Falk, 2014; Siitonen & Ranius, 2015). This means that veteran trees amongst wood pasture and parklands (including in urban areas) may be particularly suitable (Bergmeier & Roellig, 2014; Harding & Rose, 1986; Ramírez-Hernández et al., 2014; Jonsell, 2012; Jørgensen & Quelch, 2014), though this is not at all a steadfast rule as species may also be found abundantly in (perhaps more open) woodland, and particularly where there are large amounts of veteran trees and deadwood – around 60 cubic metres per hectare, according to Müller et al. (2010). Granted, they are found particularly in older (mature to veteran) trees, including within cavities that possess wood mould, water-filled rot holes, dead bark, exposed wood, sap flows, fruiting bodies (of fungi and slime moulds), mycelia of fungi, dead branches, and dead roots (Carpaneto et al., 2010; Falk, 2014; Harding & Rose, 1986; Siitonen & Ranius, 2015; Stokland et al., 2012). Beetle species may also not necessarily associate preferentially with a species (or group of species), but with the conditions aforementioned that are present within a tree (Harding & Rose, 1986; Jonsell, 2012). At times, preferable conditions may be an infrequent as one veteran tree in every hundred (Harding & Rose, 1986).
A veteran oak tree that is of prime habitat for a variety of organisms.
Despite this, species preference is observed. For broadleaved obligates, heavier shade may be more necessary, and in such instances there is a closer affinity of the beetles with fungal mycelium. Because fungi tend to produce more mycelium in cooler and more humid conditions (though this does, of course, vary with the species), the broadleaved obligates may therefore be found normally in greater abundance where conditions are more suited to fungal growth, and their presence may thus be associated with a canopy openness of as little as 20% (Bässler et al., 2010; Müller et al., 2010). This is, of course, not a steadfast rule, and many open wood pastures may support a great abundance of saproxylic beetles (Harding & Rose, 1986).
It is also important to recognise that many species of saproxylic beetle are reliant upon particular stages of the wood decay process. For instance, species that require fresh phloem tissue will only be able to colonise briefly in the first few summers following on from the death of the phloem tissue (Falk, 2014). Other species require significantly-decayed wood in a particular micro-climate, and even of a particular tree species (Harding & Rose, 1986). There also exist intricate associations between species of fungi and saproxylic insects. Inonotus hispidus, which is usually found upon ash, is the habitat for Triplax russica and Orchesia micans, whilst the coal fungus (Daldinia concentrica), also oft found upon the deadwood of ash (Fraxinus excelsior), is the main provider of habitat for Platyrhinus resinosus (Falk, 2014). The birch polypore (Fomitopsis betulina) is also host to numerous species of Coleoptera (Harding & Rose, 1986); as is the polypore Fomitopsis pinicola (Jonsson & Nordlander, 2006; Komonen, 2003; Komonen et al., 2000). This means that these species may be found where there is a suitable population of the fungus’ host species, where sporophores are present and will likely fruit again in the future, across numerous trees, and for many years. Most beetle species rely on oak more so than other tree species however, as oak generally lives for much longer and thus provides a wider array of different micro-habitats, and possesses increased compositional complexity as a result (Harding & Rose, 1986; Siitonen & Ranius, 2015).
A fruiting body of Inonotus hispidus on apple (Malus sp.). This fungus not only creates habitat in the wood that it degrades but also is a direct habitat through its sporophore.
Therefore, the loss of suitable habitat through active management programmes (including logging, and felling trees for safety reasons in urban areas) will have a very adverse impact upon saproxylic beetles, though also certain species of moth, and even species associated with saproxylic insects, including parasitic wasps, solitary wasps (which use beetle bore holes for habitat), and predatory Coleoptera (Harding & Rose, 1986; Komonen et al., 2000). Curiously, research by Carpaneto et al. (2010) concluded that trees that were ranked as the most evidently ‘hazardous’ were host to the most saproxylic beetle species, and their removal would therefore have a drastic impact upon local populations. Similarly, fragmentation of woodland patches suitable for saproxylic populations has led to a decline in the meta-populations (Grove, 2002; Komonen et al., 2000), as has deadwood removal in a managed site itself (Gibb et al., 2006). Interestingly, though not surprisingly, ‘deadwood fragmentation’ also has an adverse impact upon saproxylic insect populations (Schiegg, 2000).
Both ants and termites also benefit from the presence of deadwood. With regards to both, nests will usually form at the base of a tree or at an area where there is at least moderate decay – enough to support a viable population (Jones et al., 2003; Shigo, 1986; Stokland et al., 2012). Ants and termites both follow CODIT (compartmentalisation of damage in trees) patterns in relation to how their nests progress, and thus their territory will increase as fungal decay propagates further into the host. Ants will not feed on the decaying wood of the host however, and will simply use the decaying site as a nesting area. Conversely, termites will feast upon decayed wood and essentially control (perhaps by slowing down) the spread of fungal decay in a manner that provides as much longevity of the host as possible for a viable nesting site (Shigo, 1986). In tropical rainforests, termites are in fact considered to be one of the principal means of wood decomposition (Mori et al., 2014), and thus the provisioning of deadwood habitat is absolutely critical. Without decaying wood within trees therefore, ants and particularly termites will lack a potential habitat, and thus where a stand is actively managed populations may be markedly reduced (Donovan et al., 2007; Eggleton et al., 1995). Of course, termites are not necessarily to be desired when they are invading the wood structure of a property, and therefore deadwood is not universally beneficial (Esenther & Beal, 1979; Morales-Ramos & Rojas, 2001) – at least, when human properties are involved.
Ecologically beneficial? Yes. Economically beneficial? No. Termites can – and do – damage timber-frames buildings, as is the case here. Source: Pestec.
The presence of deadwood may also be beneficial for ground-nesting and leaf-litter dwelling spiders, which can utilise downed woody debris (particularly pieces with only slight decay) for both nesting and foraging (Varady-Szabo & Buddle, 2006). In fact, research by Buddle (2001) suggested that such spiders may more routinely utilise downed woody material when compared to elevated woody material (dead branches and telephone poles) because of the greater array of associated micro-habitats, and particularly at certain life stages – such as during egg-laying, for females (Koch et al., 2010). Furthermore, as fallen woody debris can help to retain leaf litter (or even facilitate in the build-up leaf litter), spider populations are more abundant and more diverse in sites where such woody debris is present (Castro & Wise, 2010). Therefore, where woodlands are managed and areas are clear-cut, spider populations may be markedly reduced in terms of the diversity of species. However, generalist species may benefit from the amount of cut stumps (Pearce et al., 2004). Curiously, Koch et al. (2010) suggest that spiders may perhaps benefit from woodland clearance, because the vigorous re-growth of trees and the higher light availability to the woodland floor (promoting herbaceous plant growth) increases the abundance of potential prey. Despite this, old-growth species will suffer (Buddle & Shorthouse, 2008), and thus the population structure of spider populations may dramatically change.
Soil mites are a further group that benefit from coarse woody debris, though also from hollows and holes throughout the basal region of a tree (including water-filled cavities), and from fungal sporophores and hyphae associated with wood decay (Fashing, 1998; Johnston & Crossley, 1993). Typically, termites will use fungi and insects found within the wood as a food source, and the wood structure itself will provide for an array of niche micro-habitats that are critical at different life stages of a mite. Certain mite species are obligates that associate with coarse woody debris exclusively, and may in fact only be associated with certain species’ woody debris. Additionally, mites may utilise woody debris and hollows within trees to parasitise upon other species using the ‘resource’, with both lizards and snakes being parasitised by mites following their frequenting of such resources. Beetles may also be parasitised, though the mite in such an instance may use the beetle as a means of entry into woody debris (Norton, 1980).
It is not just deadwood that arthropods will utilise, however. Foliage, both alive and abscised, is also of use (Falk, 2014). For example, the ermine moth (Yponomeutidae) will rely upon the living foliage of a host tree as a food source, and the bird cherry ermine moth (Yponomeuta evonymella) is one example of this. During late spring, larvae will fully defoliate their host Prunus padus, before pupating, emerging, and then laying eggs upon the shoots ready for the following year (Leather & Bland, 1999). Many other moth species will, during their larval stage, also behave in such a manner and thus defoliate their host – either entirely, or in part (Herrick & Gansner, 1987). Other species may alternatively have larvae mine into the leaf and feed upon the tissues within (Thalmann et al., 2003), such as horse chestnut leaf miner (Cameraria ohridella). Flies, including the holly leaf-miner (Phytomyza ilicis), will also mine leaves in a similar fashion (Owen, 1978). Ultimately however, the same purpose is served – the insect uses the living tissues of a leaf to complete its life cycle, and fuel further generations.
Bird cherry ermine moth having defoliated an entire tree. Source: Wikimedia.
Fallen leaf litter, as briefly touched upon earlier when discussing spiders, may also be of marked benefit to many arthropods. Ants, beetles, and spiders are but three examples of groups that will utilise leaf litter as a means of habitat (Apigian et al., 2006). Beetles will, for instance, rely upon leaf litter to attract potential prey, though also to provide niche micro-climates that remain relatively stable in terms of humidity, light availability, and temperature (Haila & Niemelä, 1999). Their abundance may, according to Molnár et al., (2001) be greatest at forest edges, perhaps because prey is most abundant at these edge sites (Magura, 2002). Of course, this does not mean that edges created through artificial means will necessarily improve beetle populations, as research has shown that there are few ‘edge specialists’ and therefore populations usually will go into decline where there has been significant disturbance. Unless management mimics natural mortality events of forest trees, then constituent beetle populations may thus suffer adversely (Niemelä et al., 2007).
With regards to ants, Belshaw & Bolton (1993) suggest that management practices may not necessarily impact upon ant populations, though if there is a decline in leaf litter cover then ants associated with leaf litter presence may go into – perhaps only temporary (until leaf litter accumulations once again reach desirable levels) – decline (Woodcock et al., 2011). For example, logging within a stand may reduce leaf litter abundance for some years (Vasconcelos et al., 2000), as may (to a much lesser extent) controlled burning (Apigian et al., 2006; Vasconcelos et al., 2009), though in time (up to 10 years) leaf litter may once again reach a depth suitable to support a wide variety of ant species. However, the conversion of forest stands into plantations may be one driver behind more permanently falling ant populations (Fayle et al., 2010), as may habitat fragmentation (Carvalho & Vasconcelos, 1999) – particularly when forest patches are fragmented by vast monoculture plantations of tree or crop (Brühl et al., 2003). The conversion of Iberian wood pastures to eucalyptus plantations is one real world example of such a practice (Bergmeier & Roellig, 2014).
Also of benefit to many arthropods are nectar and pollen. Bees, beetles, butterflies, and hoverflies will, for instance, use nectar from flowers as a food source (Dick et al., 2003; Kay et al., 1984), and generally (but not always) a nectar source will lack significant specificity in terms of the insect species attracted (Karban, 2015). Despite this, different chemicals secreted by different flowers, and the toxicity of certain nectar sources to particular insects, means certain tree species may only be visited by certain insect species (Adler, 2000; Rasmont et al., 2005). Tree diversity may therefore be key to sustaining healthy insect populations (Holl, 1995), and where species may prefer to frequent herbaceous plant species the presence of a diverse woodland canopy above may still be very influential (Kitahara et al., 2008). This may be because a diverse array of woody plant species increases the diversity of herbaceous species. At times, pollen may also be a reward, as may (more rarely) a flower’s scent. Karban (2015) remarks that all are collectively dubbed as ‘floral rewards’.
References
Adler, L. (2000) The ecological significance of toxic nectar. Oikos. 91 (3). p409-420.
Apigian, K., Dahlsten, D., & Stephens, S. (2006) Fire and fire surrogate treatment effects on leaf litter arthropods in a western Sierra Nevada mixed-conifer forest. Forest Ecology and Management. 221 (1). p110-122.
Bässler, C., Müller, J., Dziock, F., & Brandl, R. (2010) Effects of resource availability and climate on the diversity of wood‐decaying fungi. Journal of Ecology. 98 (4). p822-832.
Belshaw, R. & Bolton, B. (1993) The effect of forest disturbance on the leaf litter ant fauna in Ghana. Biodiversity & Conservation. 2 (6). p656-666.
Bergmeier, E. & Roellig, M. (2014) Diversity, threats, and conservation of European wood-pastures. In Hartel, T. & Plieninger, T. (eds.) European wood-pastures in transition: A social-ecological approach. UK: Earthscan.
Brühl, C., Eltz, T., & Linsenmair, K. (2003) Size does matter–effects of tropical rainforest fragmentation on the leaf litter ant community in Sabah, Malaysia. Biodiversity & Conservation. 12 (7). p1371-1389.
Buddle, C. (2001) Spiders (Araneae) associated with downed woody material in a deciduous forest in central Alberta, Canada. Agricultural and Forest Entomology. 3 (4). p241-251.
Buddle, C. & Shorthouse, D. (2008) Effects of experimental harvesting on spider (Araneae) assemblages in boreal deciduous forests. The Canadian Entomologist. 140 (4). p437-452.
Carpaneto, G., Mazziotta, A., Coletti, G., Luiselli, L., & Audisio, P. (2010) Conflict between insect conservation and public safety: the case study of a saproxylic beetle (Osmoderma eremita) in urban parks. Journal of Insect Conservation. 14 (5). p555-565.
Carvalho, K. & Vasconcelos, H. (1999) Forest fragmentation in central Amazonia and its effects on litter-dwelling ants. Biological Conservation. 91 (2). p151-157.
Castro, A. & Wise, D. (2010) Influence of fallen coarse woody debris on the diversity and community structure of forest-floor spiders (Arachnida: Araneae). Forest Ecology and Management. 260 (12). p2088-2101.
Dick, C., Etchelecu, G., & Austerlitz, F. (2003) Pollen dispersal of tropical trees (Dinizia excelsa: Fabaceae) by native insects and African honeybees in pristine and fragmented Amazonian rainforest. Molecular Ecology. 12 (3). p753-764.
Donovan, S., Griffiths, G., Homathevi, R., & Winder, L. (2007) The spatial pattern of soil‐dwelling termites in primary and logged forest in Sabah, Malaysia. Ecological Entomology. 32 (1). p1-10.
Eggleton, P., Bignell, D., Sands, W., Waite, B., Wood, T., & Lawton, J. (1995) The species richness of termites (Isoptera) under differing levels of forest disturbance in the Mbalmayo Forest Reserve, southern Cameroon. Journal of Tropical Ecology. 11 (1). p85-98.
Esenther, G. & Beal, R. (1979) Termite control: decayed wood bait. Sociobiology. 4 (2). p215-222.
Falk, S. (2014) Wood-pastures as reservoirs for invertebrates. In Hartel, T. & Plieninger, T. (eds.) European wood-pastures in transition: A social-ecological approach. UK: Earthscan.
Fashing, N. (1998) Functional morphology as an aid in determining trophic behaviour: the placement of astigmatic mites in food webs of water-filled tree-hole communities. Experimental & Applied Acarology. 22 (8). p435-453.
Fayle, T., Turner, E., Snaddon, J., Chey, V., Chung, A., Eggleton, P., & Foster, W. (2010) Oil palm expansion into rain forest greatly reduces ant biodiversity in canopy, epiphytes and leaf-litter. Basic and Applied Ecology. 11 (4). p337-345.
Gibb, H., Pettersson, R., Hjältén, J., Hilszczański, J., Ball, J., Johansson, T., Atlegrim, O., & Danell, K. (2006) Conservation-oriented forestry and early successional saproxylic beetles: responses of functional groups to manipulated dead wood substrates. Biological Conservation. 129 (4). p437-450.
Grove, S. (2002) Saproxylic insect ecology and the sustainable management of forests. Annual Review of Ecology and Systematics. 33 (1). p1-23.
Haila, Y. & Niemelä, J. (1999) Leaf litter and the small‐scale distribution of carabid beetles (Coleoptera, Carabidae) in the boreal forest. Ecography. 22 (4). p424-435.
Harding, P. & Rose, F. (1986) Pasture-Woodlands in Lowland Britain: A review of their importance for wildlife conservation. UK: NERC.
Herrick, O. & Gansner, D. (1987) Gypsy moth on a new frontier: forest tree defoliation and mortality. Northern Journal of Applied Forestry. 4 (3). p128-133.
Holl, K. (1995) Nectar resources and their influence on butterfly communities on reclaimed coal surface mines. Restoration Ecology. 3 (2). p76-85.
Jones, D., Susilo, F., Bignell, D., Hardiwinoto, S., Gillison, A., & Eggleton, P. (2003) Termite assemblage collapse along a land‐use intensification gradient in lowland central Sumatra, Indonesia. Journal of Applied Ecology. 40 (2). p380-391.
Jonsell, M. (2012) Old park trees as habitat for saproxylic beetle species. Biodiversity and Conservation. 21 (3). p619-642.
Jonsell, M. & Nordlander, G. (2004) Host selection patterns in insects breeding in bracket fungi. Ecological Entomology. 29 (6), p697-705.
Johnston, J. & Crossley, D. (1993) The significance of coarse woody debris for the diversity of soil mites. In McMinn, J. & Crossley, D. (eds.) Proceedings of the Workshop on Coarse Woody Debris in Southern Forests: Effects on Biodiversity. General Technical Report SE-94.
Jørgensen, D. & Quelch, P. (2014) The origins and history of medieval wood-pastures. In Hartel, T. & Plieninger, T. (eds.) European wood-pastures in transition: A social-ecological approach. UK: Earthscan.
Karban, R. (2015) Plant Sensing & Communication. USA: University of Chicago Press.
Kay, Q., Lack, A., Bamber, F., & Davies, C. (1984) Differences between sexes in floral morphology, nectar production and insect visits in a dioecious species, Silene dioica. New Phytologist. 98 (3). p515-529.
Kitahara, M., Yumoto, M., & Kobayashi, T. (2008) Relationship of butterfly diversity with nectar plant species richness in and around the Aokigahara primary woodland of Mount Fuji, central Japan. Biodiversity and Conservation. 17 (11). p2713-2734.
Koch, J., Grigg, A., Gordon, R., & Majer, J. (2010) Arthropods in coarse woody debris in jarrah forest and rehabilitated bauxite mines in Western Australia. Annals of Forest Science. 67 (1). p106-115.
Komonen, A. (2003) Distribution and abundance of insect fungivores in the fruiting bodies of Fomitopsis pinicola. Annales Zoologici Fennici. 40 (6). p495-504.
Komonen, A., Penttilä, R., Lindgren, M., & Hanski, I. (2000) Forest fragmentation truncates a food chain based on an old-growth forest bracket fungus. Oikos. 90 (1). p119-126.
Leather, S. & Bland, K. (1999) Naturalists’ Handbook 27: Insects on cherry trees. UK: The Richmond Publishing Co. Ltd.
Magura, T. (2002) Carabids and forest edge: spatial pattern and edge effect. Forest Ecology and Management. 157 (1). p23-37.
Molnár, T., Magura, T., Tóthmérész, B., & Elek, Z. (2001) Ground beetles (Carabidae) and edge effect in oak-hornbeam forest and grassland transects. European Journal of Soil Biology. 37 (4). p297-300.
Morales-Ramos, J. & Rojas, M. (2001) Nutritional Ecology of the Formosan Subterranean Termite (Isoptera: Rhinotermitidae) – Feeding Response to Commercial Wood Species. Journal of Economic Entomology. 94 (2). p516-523.
Mori, S., Itoh, A., Nanami, S., Tan, S., Chong, L., & Yamakura, T. (2014) Effect of wood density and water permeability on wood decomposition rates of 32 Bornean rainforest trees. Journal of Plant Ecology. 7 (4). p356-363.
Müller, J., Noss, R., Bussler, H., & Brandl, R. (2010) Learning from a “benign neglect strategy” in a national park: Response of saproxylic beetles to dead wood accumulation. Biological Conservation. 143 (11). p2559-2569.
Norton, R. (1980) Observations on phoresy by oribatid mites (Acari: Oribatei). International Journal of Acarology. 6 (2). p121-130.
Niemelä, J., Koivula, M., & Kotze, D. (2007) The effects of forestry on carabid beetles (Coleoptera: Carabidae) in boreal forests. Journal of Insect Conservation. 11 (1). p5-18.
Owen, D. (1978) The effect of a consumer, Phytomyza ilicis, on seasonal leaf-fall in the holly, Ilex aquifolium. Oikos. 31 (2). p268-271.
Pearce, J., Venier, L., Eccles, G., Pedlar, J., & McKenney, D. (2004) Influence of habitat and microhabitat on epigeal spider (Araneae) assemblages in four stand types. Biodiversity & Conservation. 13 (7). p1305-1334.
Ramírez-Hernández, A., Micó, E., de los Ángeles Marcos-García, M., Brustel, H., & Galante, E. (2014) The “dehesa”, a key ecosystem in maintaining the diversity of Mediterranean saproxylic insects (Coleoptera and Diptera: Syrphidae). Biodiversity and Conservation. 23 (8). p2069-2086.
Rasmont, P., Regali, A., Ings, T., Lognay, G., Baudart, E., Marlier, M., Delcarte, E., Viville, P., Marot, C., Falmagne, P., & Verhaeghe, J. (2005) Analysis of pollen and nectar of Arbutus unedo as a food source for Bombus terrestris (Hymenoptera: Apidae). Journal of Economic Entomology. 98 (3). p656-663.
Schiegg, K. (2000) Are there saproxylic beetle species characteristic of high dead wood connectivity?. Ecography. 23 (5). p579-587.
Shigo, A. (1986) A New Tree Biology. USA: Shigo and Trees Associates.
Siitonen, J. & Ranius, T. (2015) The Importance of Veteran Trees for Saproxylic Insects. In Kirby, K. & Watkins, C. (eds.) Europe’s Changing Woods and Forests: From Wildwood to Managed Landscapes. UK: CABI.
Stokland, J., Siitonen, J., & Jonsson, B. (2012) Biodiversity in Dead Wood. UK: Cambridge University Press.
Thalmann, C., Freise, J., Heitland, W., & Bacher, S. (2003) Effects of defoliation by horse chestnut leafminer (Cameraria ohridella) on reproduction in Aesculus hippocastanum. Trees. 17 (5). p383-388.
Varady-Szabo, H. & Buddle, C. (2006) On the relationships between ground-dwelling spider (Araneae) assemblages and dead wood in a northern sugar maple forest. Biodiversity & Conservation. 15 (13). p4119-4141.
Vasconcelos, H., Pacheco, R., Silva, R., Vasconcelos, P., Lopes, C., Costa, A., & Bruna, E. (2009) Dynamics of the leaf-litter arthropod fauna following fire in a neotropical woodland savanna. PLoS One. 4 (11). p1-9.
Vasconcelos, H., Vilhena, J., & Caliri, G. (2000) Responses of ants to selective logging of a central Amazonian forest. Journal of Applied Ecology. 37 (3). p508-514.
Woodcock, P., Edwards, D., Fayle, T., Newton, R., Khen, C., Bottrell, S., & Hamer, K. (2011) The conservation value of South East Asia’s highly degraded forests: evidence from leaf-litter ants. Philosophical Transactions of the Royal Society of London B: Biological Sciences. 366 (1582). p3256-3264.
Trees, and more specifically groups of trees, are of significant importance to avifauna. Their provisioning of food, either directly (fruits, nuts, blossom) or indirectly (attracting insects and other types of prey), in addition to their ability to act as a nesting site, roosting site, or otherwise, makes tree presence absolutely crucial to a successful and healthy bird population. Of course, different bird species will respond favourably to different tree species and stand structures, and this – amongst other aspects – is discussed below.
As alluded to above, the structure of a woodland stand will have a marked impact upon bird species present within a site. For example, active coppice woodlands will provide habitat to bird species not frequently (if at all) found in old-growth stands or even coppice of over 11-12 years since the last cycle (Fuller & Green, 1999), though wood pastures, forest glades, and even agricultural fields bordering woodland may provide niche habitat for particular birds, of which many may be associated with grasslands and the transitional zone (ecotone) between grassland and woodland (Costa et al., 2014; Hartel et al., 2014; Hinsley et al., 2015) – including the nightingale (Luscinia megarhynchos) and the chiffchaff (Phylloscopus collybita), in the UK.
A nightingale perched upon a tree branch. Source: Wikimedia.
Stand structure will also impact upon the growth of young chicks, with some species growing better in older stands and others in younger stands (Hinsley et al., 2002). This is due to some bird species feeding up in the crown of a tree, whilst others forage near to ground level. For ground foraging birds, there will likely be a lack food sources available, where canopy closure has occurred; as will there be a lack of ground-cover for nesting (Fuller & Green, 1999). Similar conditions can however be created by grazing mammals, with deer being a notable example in the UK and North America (Gill & Fuller, 2007; McShea & Rappole, 2000). Furthermore, ground-nesting and ground-foraging birds are also more sensitive to disturbance, and therefore their presence may also be limited in high-traffic areas and locations where predators (and herbivores – including deer and other grazing animals) are found in abundance (Ford et al., 2001; Fuller, 2001; Martin & McIntyre, 2007; Schmidt & Whelan, 1999). Vehicular traffic may also be an issue, and notably when a woodland site runs adjacent to a busy road (Reijnen et al., 1995). Research has therefore suggested that established woodland sites, free of major disturbance and possessing greater structural diversity than succeeding woodlands or coppiced woodlands, will provide for a greater array of bird species (Gil-Tena et al., 2007; Hinsley et al., 2009), though even amongst structurally similar habitats the species composition of a site may have a marked impact upon bird species diversity (Arnold, 1988).
In fact, a greater mix of tree species may bolster bird diversity, as was demonstrated by Díaz (2006) when bird species in pinewoods and oakwoods were found to be lower than in a stand containing both species. By a similar token, species composition may impact upon bird species that forage amongst foliage for arthropods and other food sources. Investigations by Robinson & Holmes (1984), for instance, demonstrated that the distribution of foliage within the crown of a tree will impact upon the foraging ability of particular birds; as will, but only at times, the size (and other characteristics) of foliage. Similarly, as particular tree species will attract certain arthropods, the species composition of a stand will impact upon the constituent bird species and their abundance. Thus, a mosaic of habitats that is mainly – but not at all exclusively – mature and mixed woodland may be most preferable if seeking to attract many species of bird. Such woodland need not be extensive in canopy cover however, as wood pastures attract such an abundance of insects that insectivorous birds can be found in great abundance, assuming the land is not treated with pesticides (Ceia & Ramos, 2016).
Building upon the concept of stand structure, the presence of standing deadwood is also important for birds. Whilst cycles of management are beneficial for some species, those that rely on old-growth stands with minimal management intervention are heavily reliant upon standing deadwood as a source of habitat (Drapeau et al., 2009). Those species which nest within recently-dead snags (or dead portions of living trees), including the woodpecker (Smith, 2007) – though also many species of secondary (successional) cavity-nesting species – will far more readily be found in stands of significant age that contain tracts of large (over 30cm DBH) potential habitat (Bednarz et al., 2004; Remm et al., 2006). Granted, not all standing deadwood is equal. For example, in the forests of British Columbia, USA, woodpeckers will preferentially frequent trembling aspen (Populus tremuloides), to the point that 95% of all cavity nests are found within this species – even in spite of its limited abundance within forest stands (Martin et al., 2004). Similarly, forest edge standing deadwood may be more preferable for some cavity-nesting birds (Remm et al., 2006), and at times standing deadwood created through recent forest fires may be most suitable (Nappi & Drapeau, 2011; Saab et al., 2004). Therefore, post-fire salvage logging may be detrimental to cavity-nesting birds (Hutto & Gallo, 2006). It should however be noted that not all cavity-nesting birds will create their own cavities from sites of decaying wood, and may instead use natural cavities that have formed at the branch junctions of snags (Remm et al., 2006).
A parakeet making this cavity within a large branch of London plane its nesting site. Source: Authoor, 2017.
The benefits of standing deadwood extend beyond the mere provisioning of viable nesting sites, however. They also act as suitable feeding platforms for many bird species, again including the woodpecker. In particular, decaying snags with lower wood densities will provide the suitable conditions for foraging (Farris et al., 2004; Weikel & Hayes, 1999). This is because such decaying snags attract saproxylic insects, which are viable sources of food for birds (Drapeau et al., 2009). However, this does not necessarily mean that such snags should be extensively degraded, as research has also suggested that snags with only some deterioration (through fungal decay and fire damage) are optimal for foraging (Nappi et al., 2003; Nappi et al., 2010). Without question, larger snags will normally provide for greater foraging potential, and not only because of the greater diversity of foraging site types (small branches, large branches, and the stem), but also because of the greater surface area upon which birds (including woodpeckers) may forage (Smith, 2007). By a similar token, snags can also be used for perching and communicating (Lohr et al., 2002), which could be of advantage to predatory birds and breeding birds, respectively.
Coarse woody debris (fallen deadwood) upon the woodland floor can also be of use to bird species. Lohr et al. (2002) identify such downed woody debris as being important for foraging, perching, and communicating; albeit at a generally lesser rate than standing deadwood (snags), though not always (Spetich et al., 1999). Understorey bird species may also utilise downed stems for nesting. Where coarse woody debris is removed therefore, bird species diversity and population abundance will almost certainly suffer (Riffell et al., 2011).
A bird that has used these Ganoderma brackets, which themselves reside between two buttress root of horse chestnut, as a nesting site. Source: Author, 2016.
Of course, it is not only standing (snags) and fallen deadwood (coarse woody debris) that are of benefit, but also the decaying wood of living trees. Typically, it will be trees with more extensive internal decay and thus thinner strips of functional sapwood that will be more preferable to cavity-nesting birds (Losin et al., 2006). However, it is the larger individuals within a stand that will again be more readily frequented, with research by Conner et al. (1994) finding that the red-cockaded woodpecker (Leuconotopicus borealis) requires decaying heartwood of 15cm in diameter (or greater) to form a viable nesting site. Such extensive and suitable heartwood can usually only be found in older trees (Hooper et al., 1991), which therefore outlines the importance of conserving old-growth stands and retaining mature individuals during harvesting operations. In fact, red-cockaded woodpeckers will seek-out older trees wherever possible, because of the greater heartwood extent found within such trees (Rudolph & Conner, 1991).
Furthermore, akin to standing deadwood, not all trees are equal in their provisioning of viable habitat for cavity-nesting birds. Certain bird species may favour particular trees that are being decayed by specific heart-rotting fungi. Using the red-cockaded woodpecker as an example again, it is understood that Pinus spp. being decayed by the heart rot fungus Porodaedalea pini (syn: Phellinus pini) are highly desirable sites for nesting for the species (Jackson & Jackson, 2004). Similarly, the great spotted woodpecker (Dendrocopos major) will commonly frequent large oaks complete with large tracts of decaying heartwood and fungal sporophores (Pasinelli, 2007).
Birds may also utilise the tree’s flower (florivore), fruit (frugivore), and seed crops (granivore), as a source of food. In fact, birds are considered the most significant dispersal agent of a tree’s fruit and seed crops, which is testament to the important relationship birds and trees have in this regard (Howe & Primack, 1975; Sedgley & Griffin, 1989). Certain birds are even associated largely with specific tree species, such as how the Eurasian jay’s (Garrulus glandarius) main food source is the acorn of the oak (Quercus spp.) (Vera, 2000). Open-grown mature trees may typically harbour the greatest crops (Green, 2007), and parklands, pastures (Galindo-González et al., 2000), savannas (Dean et al., 1999), and even gardens and orchards (Genghini et al., 2006; Herzog et al., 2005) may be home to many such trees.
A jay proudly carrying an acorn. Source: Phil Winter.
Unfortunately, pressure on these environments, be it in the form of grazing, chemical applications (particularly in orchards), or simply human activities, has led to declines in constituent bird populations, in some instances (Bishop et al., 2000; Elliott et al., 1994; Thiollay, 2006), though historically orchards amongst extensively-grazed wood pasture were highly valuable for bird species, which would feed upon the abundance of insects (Barnes & Williamson, 2011; Oppermann, 2014). Beyond the open-grown tree however, copses, woodlands, and great vast forests all have the ability to harbour birds, courtesy of their crops. Secondary and regenerating stands may perhaps provide for the greatest abundance and diversity of food for birds, given that the greater light levels provide suitable conditions for a wider range of plant and tree species that flower and subsequently produce fruits (Martin, 1985).
Additionally, the better light conditions mean such fruiting species are likely to be healthier and produce bigger and more plentiful fruits, which is of importance to foraging birds that seek out proteins, fats and carbohydrates from tree crops (Sedgley & Griffin, 1989) and insects attracted to flowers. One example of this would be how the plentiful silver birch (Betula pendula) stands, in Belfairs Wood (Essex, UK) during the 1970s, over-masted quite significantly and consequently attracted very large numbers of redpoll and finch (Carduelis spp.), which all foraged eagerly for the seed. By-and-large, as birds will seek-out fruits and seeds that are larger than average and in healthy supply upon a tree (Foster, 1990; Wheelwright, 1993), it is perhaps not surprising that such regenerating stands are highly desirable. Granted, closed-canopy and late-successional stands also harbour tree crops (including the acorns of Quercus spp. and keys of Fraxinus spp.) that are of huge value to birds (Greig-Smith & Wilson, 1985; Koenig & Heck, 1988). However, the poor soils (nutritionally and hydrologically) of many mature woodlands adjacent to agricultural landscapes had led to – at least in Australia – declines in fruit and seed crops and, as a result, bird population density (Watson, 2011).
Moving away from the woodland and forest stands, though not entirely returning to open-grown trees, we can observe how trees within field hedgerows can be of huge benefit to birds, as can trees within agricultural windbreaks. Benefit may come in the form of landscape connectivity, where hedgerows and windbreaks act as corridors connecting woodland patches to one-another (Davies & Pullin, 2007; Harvey, 2000; Leon & Harvey, 2006; Morelli, 2013), though they may also be used – albeit perhaps less frequently now, courtesy of increased hedgerow management (at least, in the UK) – as nesting sites and foraging sites (Benton et al., 2003; Netwon, 2004). Grass buffers either side of the hedgerow may aid with suitability for birds, as may the presence of a greater number of large trees within a hedgerow (Hinsley & Bellamy, 2000; Herzog et al., 2005).
Within urban environments, the presence of trees and hedgerows adjacent to busy roads can however have a negative impact upon birds, by increasing mortality rates (usually associated with birds flying out into oncoming traffic). Research by Orłowski (2008) concludes as such. Of course, the presence of trees is also of benefit, much like within farmland hedgerows. Urban street trees, and also those within gardens, can improve landscape connectivity, allowing for bird species to travel between more significant areas of tree cover found in parklands and urban woodlands (Sanesi et al., 2009). In particular, connectivity to older parks with remnant woodland fragments will support a greater diversity of bird species (Fernández‐Juricic, 2000). The advent of large coniferous tree (and hedge) planting in many urban areas, courtesy of the planting of the cypress and other conifers (including Chamaecyparia lawsoniana, Cupressus macrocarpa, and x Cupressocyparis leylandii), has also led to an increase in resident bird populations and primarily because of the over-winter shelter such coniferous tree species provide (Jokimäki & Suhonen, 1998; Melles et al., 2003; Rutz, 2008; Savard et al., 2000). Furthermore, sheltered trees within the urban landscape that have abundant fruit and seed crops can be of huge benefit to birds, by providing essential food sources in an otherwise somewhat undesirable landscape. For such reasons, urban parks and woodlands may potentially provide the best conditions for certain feeding birds, though large gardens complete with dense vegetation may also be of great importance. Tree-lined streets may also be critical, and notably so if trees are large, have dense crowns, and have an edible fruit or seed crop.
Large leyland cypress specimens inter-planted with poplar cultivars offer suitable nesting sites in this harsh industrial zone. Source: Author, 2016.
References
Arnold, G. (1988) The Effects of Habitat Structure and Floristics on the Densities of Bird Species in Wandoo Woodland. Wildlife Research. 15 (5). p499-510.
Barnes, G. & Williamson, T. (2011) Ancient Trees in the Landscape: Norfolk’s arboreal heritage. UK: Windgather Press.
Bednarz, J., Ripper, D., & Radley, P. (2004) Emerging concepts and research directions in the study of cavity-nesting birds: keystone ecological processes. The Condor. 106 (1). p1-4.
Benton, T., Vickery, J., & Wilson, J. (2003) Farmland biodiversity: is habitat heterogeneity the key?. Trends in Ecology & Evolution. 18 (4). p182-188.
Bishop, C., Ng, P., Mineau, P., Quinn, J., & Struger, J. (2000) Effects of pesticide spraying on chick growth, behavior, and parental care in tree swallows (Tachycineta bicolor) nesting in an apple orchard in Ontario, Canada. Environmental Toxicology and Chemistry. 19 (9). p2286-2297.
Ceia, R. & Ramos, J. (2016) Birds as predators of cork and holm oak pests. Agroforestry Systems. 90 (1). p159-176.
Costa, A., Madeira, M., Santos, J., & Plieninger, T. (2014) Recent dynamics of evergreen oak wood-pastures in south-western Iberia. In Hartel, T. & Plieninger, T. (eds.) European wood-pastures in transition: A social-ecological approach. UK: Earthscan.
Davies, Z. & Pullin, A. (2007) Are hedgerows effective corridors between fragments of woodland habitat? An evidence-based approach. Landscape Ecology. 22 (3). p333-351.
Dean, W., Milton, S., & Jeltsch, F. (1999) Large trees, fertile islands, and birds in arid savanna. Journal of Arid Environments. 41 (1). p61-78.
Díaz, L. (2006) Influences of forest type and forest structure on bird communities in oak and pine woodlands in Spain. Forest Ecology and Management. 223 (1). p54-65.
Drapeau, P., Nappi, A., Imbeau, L., & Saint-Germain, M. (2009) Standing deadwood for keystone bird species in the eastern boreal forest: managing for snag dynamics. The Forestry Chronicle. 85 (2). p227-234.
Elliott, J., Martin, P., Arnold, T., & Sinclair, P. (1994) Organochlorines and reproductive success of birds in orchard and non-orchard areas of central British Columbia, Canada, 1990–91. Archives of Environmental Contamination and Toxicology. 26 (4). p435-443.
Farris, K., Huss, M., & Zack, S. (2004) The role of foraging woodpeckers in the decomposition of ponderosa pine snags. The Condor. 106 (1). p50-59.
Fernández‐Juricic, E. (2000) Bird community composition patterns in urban parks of Madrid: the role of age, size and isolation. Ecological Research. 15 (4). p373-383.
Ford, H., Barrett, G., Saunders, D., & Recher, H. (2001) Why have birds in the woodlands of Southern Australia declined?. Biological Conservation. 97 (1). p71-88.
Foster, M. (1990) Factors influencing bird foraging preferences among conspecific fruit trees. The Condor. 92 (4). p844-854.
Fuller, R. (2001) Responses of woodland birds to increasing numbers of deer: a review of evidence and mechanisms. Forestry. 74 (3). p289-298.
Fuller, R. & Green, G. (1998) Effects of woodland structure on breeding bird populations in stands of coppiced lime (Tilia cordata) in western England over a 10-year period. Forestry. 71 (3). p199-218.
Galindo‐González, J., Guevara, S., & Sosa, V. (2000) Bat‐and bird‐generated seed rains at isolated trees in pastures in a tropical rainforest. Conservation Biology. 14 (6). p1693-1703.
Genghini, M., Gellini, S., & Gustin, M. (2006) Organic and integrated agriculture: the effects on bird communities in orchard farms in northern Italy. Biodiversity & Conservation. 15 (9). p3077-3094.
Gil-Tena, A., Saura, S., & Brotons, L. (2007) Effects of forest composition and structure on bird species richness in a Mediterranean context: implications for forest ecosystem management. Forest Ecology and Management. 242 (2). p470-476.
Gill, R. & Fuller, R. (2007) The effects of deer browsing on woodland structure and songbirds in lowland Britain. Ibis. 149 (2). p119-127.
Greig-Smith, P. & Wilson, M. (1985) Influences of seed size, nutrient composition and phenolic content on the preferences of bullfinches feeding in ash trees. Oikos. 44 (1). p47-54.
Green, T. (2007) Stating the obvious: the biodiversity of an open-grown tree – from acorn to ancient. In Rotherham, I. (ed.) The History, Ecology, and Archaeology of Medieval Parks and Parklands. UK: Wildtrack Publishing.
Hartel, T., Hanspach, J., Abson, D., Máthé, O., Moga, C., & Fischer, J. (2014) Bird communities in traditional wood-pastures with changing management in Eastern Europe. Basic and Applied Ecology. 15 (5). p385-395.
Harvey, C. (2000) Colonization of agricultural windbreaks by forest trees: effects of connectivity and remnant trees. Ecological Applications. 10 (6). p1762-1773.
Herzog, F., Dreier, S., Hofer, G., Marfurt, C., Schüpbach, B., Spiess, M., & Walter, T. (2005) Effect of ecological compensation areas on floristic and breeding bird diversity in Swiss agricultural landscapes. Agriculture, Ecosystems & Environment. 108 (3). p189-204.
Hinsley, S. & Bellamy, P. (2000) The influence of hedge structure, management and landscape context on the value of hedgerows to birds: a review. Journal of Environmental Management. 60 (1). p33-49.
Hinsley, S., Hill, R., Fuller, R., Bellamy, P., & Rothery, P. (2009) Bird species distributions across woodland canopy structure gradients. Community Ecology. 10 (1). p99-110.
Hinsley, S., Fuller, R., & Ferns, P. (2015) The Changing Fortunes of Woodland Birds in Temperate Europe. In Kirby, K. & Watkins, C. (eds.) Europe’s Changing Woods and Forests: From Wildwood to Managed Landscapes. UK: CABI.
Hooper, R., Lennartz, M., & Muse, H. (1991) Heart rot and cavity tree selection by red-cockaded woodpeckers. The Journal of Wildlife Management. 55 (2). p323-327.
Howe, H. & Primack, R. (1975) Differential seed dispersal by birds of the tree Casearia nitida (Flacourtiaceae). Biotropica. 7 (4). p278-283.
Hutto, R. & Gallo, S. (2006) The effects of postfire salvage logging on cavity-nesting birds. The Condor. 108 (4). p817-831.
Jackson, J. & Jackson, B. (2004) Ecological relationships between fungi and woodpecker cavity sites. The Condor. 106 (1). p37-49.
Jokimäki, J. & Suhonen, J. (1998) Distribution and habitat selection of wintering birds in urban environments. Landscape and Urban Planning. 39 (4). p253-263.
Koenig, W. & Heck, M. (1988) Ability of two species of oak woodland birds to subsist on acorns. The Condor. 90 (3). p705-708.
Leon, M. & Harvey, C. (2006) Live fences and landscape connectivity in a neotropical agricultural landscape. Agroforestry Systems. 68 (1). p15-26.
Lohr, S., Gauthreaux, S., & Kilgo, J. (2002) Importance of coarse woody debris to avian communities in loblolly pine forests. Conservation Biology. 16 (3). p767-777.
Losin, N., Floyd, C., Schweitzer, T., & Keller, S. (2006) Relationship between aspen heartwood rot and the location of cavity excavation by a primary cavity-nester, the Red-naped Sapsucker. The Condor. 108 (3). p706-710.
Martin, K., Aitken, K, & Wiebe, K. (2004) Nest sites and nest webs for cavity-nesting communities in interior British Columbia, Canada: nest characteristics and niche partitioning. The Condor. 106 (1). p5-19.
Martin, T. (1985) Selection of second-growth woodlands by frugivorous migrating birds in Panama: an effect of fruit size and plant density?. Journal of Tropical Ecology. 1 (2). p157-170.
Martin, T. & McIntyre, S. (2007) Impacts of livestock grazing and tree clearing on birds of woodland and riparian habitats. Conservation Biology. 21 (2). p504-514.
McShea, W. & Rappole, J. (2000) Managing the abundance and diversity of breeding bird populations through manipulation of deer populations. Conservation Biology. 14 (4). p1161-1170.
Melles, S., Glenn, S. and Martin, K., 2003. Urban bird diversity and landscape complexity: species-environment associations along a multiscale habitat gradient. Conservation Ecology. 7 (1). p1-22.
Morelli, F. (2013) Relative importance of marginal vegetation (shrubs, hedgerows, isolated trees) surrogate of HNV farmland for bird species distribution in Central Italy. Ecological Engineering. 57 (1). p261-266.
Nappi, A. & Drapeau, P. (2011) Pre-fire forest conditions and fire severity as determinants of the quality of burned forests for deadwood-dependent species: the case of the black-backed woodpecker. Canadian Journal of Forest Research. 41 (5). p994-1003.
Nappi, A., Drapeau, P., Giroux, J., & Savard, J. (2003) Snag use by foraging black-backed woodpeckers (Picoides arcticus) in a recently burned eastern boreal forest. The Auk. 120 (2). p505-511.
Nappi, A., Drapeau, P., Saint-Germain, M., & Angers, V. (2010) Effect of fire severity on long-term occupancy of burned boreal conifer forests by saproxylic insects and wood-foraging birds. International Journal of Wildland Fire. 19 (4). p500-511.
Newton, I. (2004) The recent declines of farmland bird populations in Britain: an appraisal of causal factors and conservation actions. Ibis. 146 (4). p579-600.
Oppermann, R. (2014) Wood-pastures as examples of European high nature value landscapes. In Hartel, T. & Plieninger, T. (eds.) European wood-pastures in transition: A social-ecological approach. UK: Earthscan.
Orłowski, G. (2008) Roadside hedgerows and trees as factors increasing road mortality of birds: implications for management of roadside vegetation in rural landscapes. Landscape and Urban Planning. 86 (2). p153-161.
Pasinelli, G. (2007) Nest site selection in middle and great spotted woodpeckers Dendrocopos medius & D. major: implications for forest management and conservation. Biodiversity and Conservation. 16 (4). p1283-1298.
Reijnen, R., Foppen, R., Braak, C., & Thissen, J. (1995) The effects of car traffic on breeding bird populations in woodland. III. Reduction of density in relation to the proximity of main roads. Journal of Applied Ecology. 32 (1). p187-202.
Remm, J., Lohmus, A., & Remm, K. (2006) Tree cavities in riverine forests: What determines their occurrence and use by hole-nesting passerines?. Forest Ecology and Management. 221 (1). p267-277.
Riffell, S., Verschuyl, J., Miller, D., & Wigley, T. (2011) Biofuel harvests, coarse woody debris, and biodiversity–a meta-analysis. Forest Ecology and Management. 261 (4). p878-887.
Robinson, S. & Holmes, R. (1984) Effects of plant species and foliage structure on the foraging behavior of forest birds. The Auk. 101 (4). p672-684.
Rudolph, D. & Conner, R. (1991) Cavity tree selection by red-cockaded woodpeckers in relation to tree age. The Wilson Bulletin. 103 (3). p458-467.
Rutz, C. (2008) The establishment of an urban bird population. Journal of Animal Ecology. 77 (5). p1008-1019.
Saab, V., Dudley, J., & Thompson, W. (2004) Factors influencing occupancy of nest cavities in recently burned forests. The Condor. 106 (1). p20-36.
Sanesi, G., Padoa-Schioppa, E., Lorusso, L., Bottoni, L., & Lafortezza, R. (2009) Avian ecological diversity as an indicator of urban forest functionality. Results from two case studies in Northern and southern Italy. Journal of Arboriculture. 35 (2). p80-86.
Savard, J., Clergeau, P., & Mennechez, G. (2000) Biodiversity concepts and urban ecosystems. Landscape and Urban Planning. 48 (3). p131-142.
Schmidt, K. & Whelan, C. (1999) Nest predation on woodland songbirds: when is nest predation density dependent?. Oikos. 87 (1). p65-74.
Sedgley, M. & Griffin, A. (1989) Sexual Reproduction of Tree Crops. UK: Academic Press.
Smith, K. (2007) The utilization of dead wood resources by woodpeckers in Britain. Ibis. 149 (2). p183-192.
Spetich, M., Shifley, S., & Parker, G. (1999) Regional distribution and dynamics of coarse woody debris in Midwestern old-growth forests. Forest Science. 45 (2). p302-313.
Thiollay, J. (2006) Large bird declines with increasing human pressure in savanna woodlands (Burkina Faso). Biodiversity & Conservation. 15 (7). p2085-2108.
Vera, F. (2000) Grazing Ecology and Forest History. UK: CABI Publishing.
Watson, D. (2011) A productivity-based explanation for woodland bird declines: poorer soils yield less food. Emu. 111 (1). p10-18.
Weikel, J. & Hayes, J. (1999) The foraging ecology of cavity-nesting birds in young forests of the northern coast range of Oregon. The Condor. 101 (1). p58-66.
Wheelwright, N. (1993) Fruit size in a tropical tree species: variation, preference by birds, and heritability. Vegetatio. 107 (1). p163-174.
The extent of attention as to exactly how critical trees are for fish populations is unfortunately not all that significant (in comparison to the study or trees and birds, for example), though this is not necessarily surprising – this is perhaps because fish spend their lives largely under water, and thus their presence is not necessarily recognised to the degree it would be if fish were land-based organisms. However, there is certainly a healthy array of research that has been undertaken into this relationship of trees and fish within the forest ecosystem, as is demonstrated below.
Many undisturbed pools (areas of slow-moving or still water within in rivers and streams) in forests are either created or enhanced by the presence of deadwood (as either driftwood or sunken wood). Such deadwood presence can also raise water levels locally and create a diverse range of aquatic habitats (Hodge & Peterken, 1998) by damming up rivers and streams, and reducing flow velocity (Barbour et al., 2001; Gippel et al., 1996). Large woody debris (including fallen stems and large branches) is particularly critical in this regard, and research has shown that nearly 30% of pools within a stream or river may be created by such woody debris (Mossop & Bradford, 2004).
A gathering of many fallen branches significantly obstructs the flow of this stream through the New Forest, UK. Such obstruction creates niche habitats on both sides of the log jam. Source: Author (2016).
Other research has, whilst not focussing on large woody debris exclusively, identified that as much as 75% of all pools may be created from submerged woody debris (Robison & Beschta, 1990a). Through the creation of these habitats, fish populations can increase, as their range of viable habitat increases – notably for feeding and spawning (Harvey, 1998). However, because even the largest of woody debris will likely not persist for over 50 years, there is a need for a continuous replenishment if streams and rivers are to retain the presence of deadwood-induced pools (Hyatt & Naiman, 2001). When pools are instead created by wood jams, which are made of small (and sometimes also large) branches and stems clustered together, their average viable retention time may only be between 2-3 years (Lisle, 1986). Again, a need for a constant supply of such deadwood is necessary, and this should obviously mean management practices retain trees that can constantly provide for such woody material (Robison & Beschta, 1990b).
Driftwood may be particularly beneficial for fish populations, as not only will its presence control flow velocity, but also protect its banks from erosion, create waterfalls and pools, and thus provide protection for fish spawning as well as increasing habitat diversity (Gurnell et al., 2002). Additionally, driftwood can provide hiding places for species of fish, assisting either in their predatory pursuits or in evading predation (Crook & Robertson, 1999; Werneyer & Kramer, 2005).
Sunken (or partially submerged) deadwood, for those fish species which are insectivorous, can also be highly valuable (Barbour et al., 2001). The wood’s provision of habitat for invertebrates means there is a potential abundance of prey for such insectivorous fish (O’Connor, 1992). A study into the effects of deforestation on wood input levels into woodland stream environments there unsurprisingly showed how reduced amounts of sunken deadwood led to reduced fish diversity and abundance (Wright & Flecker, 2004). In such wood-void streams, wood-eating fish (such as certain species of catfish, whilst not ‘true’ xylivores) may also suffer (German & Bittong, 2009; Lujan et al., 2011), though the loss of diversity in a stream (or river) environment, both because of reduced wood presence and the faster flow associated with such a lack of wood, may also have wider implications for fish species overall (Lancaster et al., 2001; Shields & Smith, 2002; Tsui et al., 2000); particularly when it is understood that a lack of (large) sunken wood is indicative of a degraded stream (Shields et al., 2006). It is also suggested that sunken wood may aid with orientation for fish (Crook & Robertson, 1999).
Some significantly-decayed deadwood from a fallen willow (Salix sp.) will offer aquatic organisms – including fish – the opportunity to forage and seek shelter. Source: Author (2016).
Deadwood that has fallen and become (partially) submerged is also beneficial, as previously ascertained, because it creates pools within a stream or river ecosystem. These pools are areas of a stream or river where the flow is potentially very slow, and in the redwood forests of California downed trunks and branches of trees are considered to be crucial for constituent salmon populations (Barbour et al., 2001). Notably, in areas of steeper ground, this fallen deadwood can create tiers of pools, which actually enable salmon (that travel upstream to breed) to ascend up the river with more ease, as the salmon can ‘leap’ from one pool to another, and swim against a current with reduced velocity (which is critical for the enabling of salmon to conserve vital energy). These pools also reduce bankside erosion and catch up to 85% of sediment (which may amass behind a large branch or stem, though perhaps even more significantly amongst larger wood jams comprised of deadwood of varying sizes), ensuring the rate of sedimentation of the stream or river is slow and sustainable (Berg et al., 1998; Smith et al., 1993; Thevenet et al., 1998). This is important for the salmon, as females nest within the clean gravel beds in the riverbed, and any marked rate of sedimentation would prohibit this (Madej & Ozaki, 2009). These nesting sites may also, in fact, be located within close proximity to large pieces of woody debris (Senter & Pasternack, 2011). The very same deadwood can also support plant life, particularly when a large stem has fallen across a river, and therefore the plants growing atop the log can shade the river and keep the water cooler – this is also critical for the salmon, which prefer cooler waters (Welsh et al., 2001).
This willow has fallen but remains alive, offering a further and somewhat different aspect to the aquatic environment. Source: Author (2016).
Across the United States, in the Appalachian Mountains, research by Jones et al. (1999) has also revealed that the reduction in sedimentation created by fallen woody debris is critical for other species of fish (including the rainbow trout Oncorhynchus mykiss), that spawn in sediment-free riffles within the forest areas of the mountains. Furthermore, their research highlighted that deforestation along riparian zones as little as 1km in length can have massive adverse effects upon the quality of habitat for fish, due to the removal of the source of such critical deadwood. The associated re-growth after the felling, whilst still injecting debris into the water courses, cannot match the size of the debris from older-growth stands, and therefore rainbow trout occur less frequently and at lesser densities (Flebbe & Dolloff, 1995). Deforestation also increases the risk of severe flooding and high flow velocity within the Appalachian Mountains, which can both extensively decimate viable habitat for rainbow trout within the ecosystem. In part, this is because such factors eliminate the fauna that occupy the river bed, which the trout almost exclusively predate upon.
Beyond the realm of deadwood, the beneficial impacts of shading by large trees adjacent to such aquatic environments can also improve the suitability of the habitat for fish (Beschta, 1997; Larson & Larson, 1996). Using the redwood forests as an example once again, it has been recognised that large conifers that reside by a water course cast shade and thus reduce maximum temperatures and the risk of thermal pollution (Madej et al., 2006). Such cooler temperatures, much like how deadwood can support plants that shade and cool waters, protects critical nesting locations for female salmon, reduces the subsequent mortality of juvenile salmon, and improves their growth rate.
The shade this single hornbeam (Carpinus betulus) provides the river beneath, whilst not necessarily significant, will be of measurable benefit. Source: Author (2016).
Beyond California, the cooler waters created through significant (50-80%) canopy shading are equally as important for fish, for similar reasons (Broadmeadow & Nisbet, 2004; Broadmeadow et al., 2011; Swift Jr & Messer, 1971). Such canopy shade may also enable for rivers and streams to support macrophytes (plants growing in or near water), which can act as a food source for some fish species both directly and indirectly. Similarly, they can provide refuge for fish seeking shelter from predators (Pusey & Arthington, 2003). Therefore, retaining riparian trees is mandatory, if viable habitats for fish are to be protected (Young, 2000).
A line of willow and ash (Fraxinus excelsior) dresses the southern side of this river, meaning the water remains continually shaded throughout the day. Source: Author (2016).